АНАЛИЗ ФЛОРЫ ОСТРОВОВ
Не вызывает сомнений, что островные флоры Северной Охотии во флори-стическом
отношении несамостоятельны и представляют собой часть более крупной флористической
единицы - Прибрежно-Охотского флористического района. Известны только 7
видов, найденных на островах и не обнаруженных (пока?) на ближайшем
материковом побережье. Местонахождения этих видов на островах Северной
Охотии являются реликтовыми, поскольку их основные современные ареалы удалены,
порой на значительное расстояние, к северу (Puccinellia vaginata, Calamagrostis
holmii, Taraxacum anadyricum, Taraxacum tamarae), востоку (Chrysosplenium
rimosum) и югу (Glyceria alnasteretum, Saxifraga idzuroei) от рассматриваемой
территории.
Побережье Северной Охотии в очерченных нами границах занимает дос-таточно
большую площадь и флора его является, конечно, не конкретной, а сборной
флорой в понимании А.И.Толмачева (1986). Однако, точное установле-ние ботанико-географических
рубежей, проходящих по территории Прибрежно-Охотского флористического района
требует более плотного обследования. Пока методом конкретных (локальных)
флор были изучены только 5 участков на п-ове Кони и в его окрестностях
(Мочалова, 1997).
При анализе флоры мы ставили перед собой задачу показать, какие виды
и группы видов принимают участие в сложении островных флор Северной Охо-тии,
а какие, напротив, там не встречаются. Задача эта решалась сравнением флоры
всего побережья с составной частью этой флоры - островной, с детали-зацией
до весьма мелких естественных подразделений островной флоры. Для сравнения
была использована также флора п-ова Кони, как территории, проме-жуточной
по своему положению и природно-климатическим условиям между островной и
типично материковой.
Флора островов Тауйской губы - “собирательный образ”, который являет-ся,
тем не менее, реальной совокупностью видов, обитающих на островах, рас-положенных
в Тауйской губе. На основе анализа сводного списка можно судить об общих
закономерностях структуры островных флор по сравнению с флорой материкового
побережья, а особенности выявляются при рассмотрении флоры каждого острова.
Флора Ямских о-вов изучена недостаточно; ее отличия от флоры о-вов Тауйской
губы рассматриваются на примере флоры о-ва Маты-киль.
5.1. Оценка уровня таксономического разнообразия
Уровень видового богатства природной флоры побережья Северной
Охо-тии, п-ова Кони в целом и его западной части , островов Тауйской
губы (объе-диненный список и 8 островных флор, расположенных в порядке
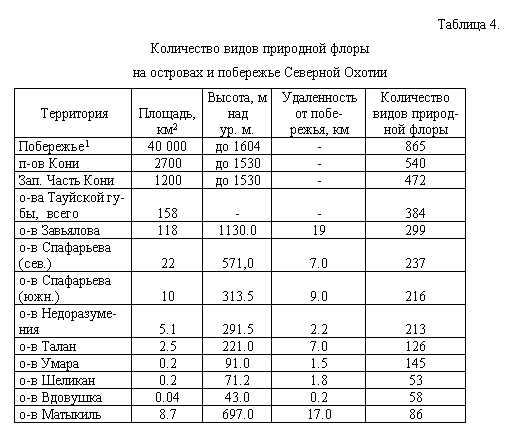
уменьшения площади), а также о-ва Матыкиль представлен в табл. 4.
Флора Прибрежно-Охотского флористического района представлена на 63%
во флоре п-ова Кони, на 55% во флоре западной части п-ова Кони, на
45% во флоре всех островов Тауйской губы, с закономерным снижением этого
пока-зателя до 6-35% в зависимости от особенностей каждого острова. Рассмотрим
факторы, от которых зависит конкретное количество видов, обитающих на том
или ином острове.
Со времени Скоу и Декандоля известно, что количество видов во флоре
зависит от размера территории, географической широты и сухости климата,
при этом флора горных стран богаче, нежели соседних равнинных (цит. по:
Малы-шев, 1969). Предпринимались попытки выразить эту зависимость математиче-ски
с учетом одного (как правило, площади) или нескольких факторов, обзор литературы
по этому вопросу дан в работах Л.И.Малышева (1969, 1972) и Р.Бешела (1969).
Для острова, кроме того, имеет значение удаленность от ма-терика
и от других островов, особенно заметно проявляющаяся в обеднении флоры
изолированных океанических островов южного полушария (Толмачев, 1970, 1986).
Очевидно, что из перечисленных абиотических факторов для ост-ровов Северной
Охотии наибольшее значение имеет размер острова. Прежде, чем перейти
к обсуждению этой зависимости, рассмотрим основные представ-ления островной
экологии и возможность их применения для наших целей.
Согласно теории динамического равновесия (MacArthur, Wilson, 1963, 1967)
число видов в изоляте (на острове) является результатом равновесия про-цессов
их иммиграции и локального выпадения. Стало аксиомой, что скорость
колонизации и скорость вымирания видов зависят от площади изолята и его
удаленности от источника распространения диаспор. Представляется достаточ-но
очевидным, что чем меньше размер острова и чем дальше он удален от побе-режья,
тем легче нарушается равновесие между процессами локального выпа-дения
видов и их иммиграции, и в результате снижается уровень видового раз-нообразия.
Такое сокращение видового богатства известно под названием “ост-ровной
эффект” (Бигон и др., 1989).
Несмотря на то, что теория динамического равновесия R. MacArtur и E.Wilson
(1963, 1967), предсказывающая островной эффект, стимулировала зна-чительное
число исследований (Nilsson, Nilsson, 1982, 1983; Preston, 1962; Westman,
1983 и др.) , вопрос - существует ли он как таковой, или же острова беднее
видами из-за своей малой территории (теория случайного распределения видов;
Coleman, 1981), небольшого набора местообитаний (Connor, McCoy, 1979) или
других факторов - все еще остается без ответа (Connor, Simberloff, 1978;
Симберлофф, 1982, 1988; Бигон и др., 1989; Kelly et al., 1989). Наиболее
часто для тестирования теории равновесия используется установление
корре-ляции между числом видов, площадью и удаленностью изолята от источника
распространения диаспор. Совершенствование этого теста шло по пути перехо-да
от анализа общего числа видов, обитающих на островах, к анализу варьиро-вания
числа видов, обитающих на определенных типах местообитаний и далее - в
пределах фиксированной площади определенных типов местообитаний. Од-нако,
во многих случаях отделение эффектов "число видов - площадь" и "число видов
- расстояние" от влияния параметров среды, по которым отличается один изолят
от другого, становится практически невозможным. В качестве способа, который
позволяет исключить “шумовое влияние” среды, маскирующее ост-ровной
эффект, предлагается анализ степени флористической неполночленно-сти фитоценозов
(Акатов, 1995).
Рассматривая возможности применения методов островной биогеографии
к анализу флоры островов Северной Охотии, мы пришли к выводу, что выявле-ние
и количественное измерение островного эффекта (который нужно понимать как
генетические процессы в изолированных популяциях, ведущие к уменьше-нию
видового разнообразия сообществ), представляет собой сложную теорети-ческую
проблему, поиск путей разрешения которой выходит за рамки нашего исследования.
Для подтверждения этих рассуждений обратимся опять к табл. 4. На о-ве
Завьялова найдены 299 видов, и размер его (118 км2) почти соответствует
ареал-минимуму конкретной флоры, который составляет 100 км2 (Толмачев,
1931, 1986 и др.). Если сравнить уровень видового богатства о-ва Завьялова
и локаль-ных флор п-ова Кони (Мочалова, 1997), насчитывающих 322-380 видов,
можно сделать вывод об относительной бедности островных флор по сравнению
со сходными по площади участками материкового побережья, что в целом не
вы-зывает сомнений. Тем не менее, вывод этот не будет достаточно
адекватным, поскольку сравниваемые территории заведомо неравнозначны, и
локальные флоры п-ова Кони, видимо, одни из наиболее богатых в Северной
Охотии. Как будет показано в дальнейшем, средний удельный уровень флористического
бо-гатства на побережье несколько ниже, чем на островах. Выбор участка
для сравнения, сходного по площади и набору экотопов с каким-либо островом
представляется нам утопической задачей, т.к. любой остров является уникаль-ным
и единственным в своем роде образованием. Здесь мы опять приходим
к выводу, что сравнение флористических списков с целью выявления островного
эффекта возможно на ценотическом уровне, но не путем сравнения всей флоры
острова с участком равной площади на материковом побережье.
Основная причина того, что острова Северной Охотии не являются иде-альным
объектом для тестирования теории равновесия - это, конечно, варьиро-вание
качества абиотической и биотической среды (емкости среды). Влияние
параметров среды на уровень видового богатства на каждом острове проявляет-ся
в различиях геоморфологического строения (которое определяет разнообра-зие
экотопов), степени воздействия морских ветров на растительный покров, а
также такого специфического фактора, как жизнедеятельность морских колони-альных
птиц (о-ва Талан, Шеликан, Умара и Ямские о-ва).
Попробуем показать сложность отделения эффектов "число видов - пло-щадь"
и "число видов - расстояние" от влияния других факторов, определяющих уровень
видового богатства, на конкретных примерах. Для того, чтобы выявить зависимость
между площадью острова и числом обитающих на нем видов, мы использовали
уравнение Аррениуса (Малышев, 1972):
y = axz , или log y = log a + z log x, где аргумент x - размер
площади, функция y - количество видов растений на этой площади, a - количество
видов растений на единице площади (плотность флоры), z - показатель пространст-венного
разнообразия флоры.
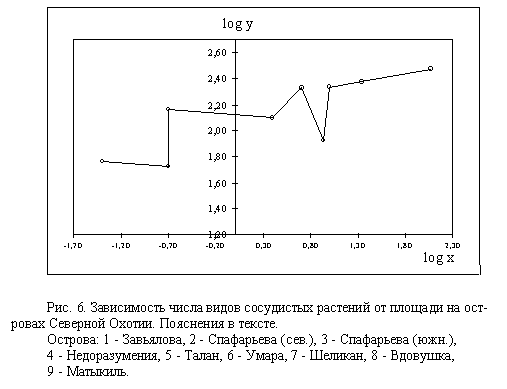
Данные по 9 островным флорам (табл. 4) были преобразованы в логариф-мический
вид и представлены в виде графика (рис. 6). На графике видно, что линейная
зависимость между log x и log y наблюдается только для о-вов Завья-лова
и Спафарьева (ОФ 1-3), и если продолжить прямую линию, то в точке
пе-ресечения с осью ординат получаем log y = log a = 2,20. Теперь мы можем
уз-нать величину z, подставляя в уравнение Аррениуса данные по о-вам Завьялова
и Спафарьева. После обратного преобразования уравнение принимает следую-щий
вид: y = 158 x 0.133 , где аргумент x - размер площади, км2, функция y
- ко-личество видов растений на этой площади, 158 - количество видов растений
на 1 км2 (плотность флоры, или удельный уровень видового богатства), 0.133
- по-казатель пространственного разнообразия флоры.
Известно, что показатель пространственного разнообразия флоры оцени-вается
величиной 0.17 на Британских о-вах, во Франции - 0.21, на побережье Калифорнии
(при учете только туземной флоры) - 0.20, в высокогорьях Восточ-ного Саяна
- 0.23, в высокогорьях Станового нагорья - 0.24, на плато Путорана - 0.11;
количество видов на 1 км2 установлено для окрестностей озера Хая-Кюель
(плато Путорана) - 173 вида, на п-ове Ямал - 92-140 видов (Малышев, 1972а,
1976; Ребристая, 1987). Учитывая эти сведения, можно заключить, что флора
о-вов Северной Охотии характеризуется относительно небольшим (но не мини-мальным)
пространственным разнообразием и плотностью. К сожалению, мы не располагаем
данными, позволяющими точно судить о плотности флоры (на 1 км2) и мере
пространственного разнообразия флоры (постоянные в уравнении Аррениуса)
на побережье Северной Охотии. Установление этих показателей требует специальных,
притом скрупулезных исследований (Малышев, 1972а, 1976; Ребристая, 1987);
не вызывает также сомнений, что плотность флоры и ее пространственное разнообразие
подвержены большому варьированию под влиянием местных условий. Мы, однако,
не могли не воспользоваться имеющимися в нашем распоряжении цифрами по
Прибрежно-Охотскому флористическому району и по п-ову Кони (табл. 4), хотя
пересчет при большой экстрапо-ляции может дать искаженные результаты. Полученное
уравнение описывает фактическое распределение следующим образом:
y = 138 x 0.173, то есть количество видов на 1 км2 (плотность флоры)
на по-бережье несколько меньше, чем на островах, а пространственное разнообразие
- больше. Противоречие здесь кажущееся, поскольку на побережье в ландшафтах
преобладают флористически бедные комплексы, что в пересчете на всю терри-торию
дает меньший уровень удельного флористического богатства. По этому уравнению
можно рассчитать средний уровень видового богатства для локаль-ных флор
Северной Охотии (100 км2), он составляет 306 видов. Согласно А.И.Толмачеву
(1941, 1986), такое количество видов характерно для гипоаркти-ческих флор
(150-300 видов, иногда, вероятно, 350). Даже если учесть, что наи-более
богатые ЛФ Северной Охотии содержат около 400 видов, они уступают типичным
бореальным флорам по уровню видового богатства (который оцени-вается в
500-800 видов). В Сибири, однако, известны и относительно бедные ЛФ даже
в подзоне южной тайги, например, на юго-востоке Томской обл., на-считывающие
в среднем около 440 видов (Пяк, Зверев, 1997). То есть по уровню видового
богатства флору Северной Охотии можно отнести к переходной меж-ду гипоарктической
и бореальной.
Вернемся к вопросу об относительной бедности островных флор. Ланд-шафтная
структура островов обусловливает пестрый состав и быструю смену растительных
сообществ (то есть высокий уровень видового богатства флоры) на небольшом
пространстве, но при увеличении площади разнообразие ланд-шафтов не возрастает
(нет речных долин, болот и т.п.), следовательно, увеличи-вается площадь,
занятая однообразными сообществами, и разнообразие флоры на острове оказывается
меньшим, чем на материковом побережье. Какова же “критическая” площадь,
обеспечивающая больший уровень видового разнооб-разия флоры на островах,
по сравнению с побережьем? Если оперировать полу-ченными нами уравнениями,
описывающими зависимость количества видов от площади на островах (y = 158
x0.133 ) и побережье (y = 138 x 0.173), то получается, что на территории
меньше 25 км2 островные флоры богаче, чем “средние” ма-териковые. При пересчете
на 100 км2 по уравнению для островных флор полу-чаем цифру 292 вида,
что несколько меньше, чем по уравнению для побережья; при дальнейшем увеличении
расчетной площади разница возрастает - всецело за счет большего пространственного
разнообразия флоры, установленного для побережья. Таким образом, по расчетным
данным, уровень видового богатства флоры на всех островах Северной Охотии,
кроме о-ва Завьялова, оказывается относительно большим, чем на материковом
побережье.
Один из способов выявления островного эффекта основан на сравнении
графиков зависимости “число видов-площадь” для островов и произвольно вы-бранных
участков (выборок) материка. В биогеографических работах исполь-зуют обычно
формулу, которая отличается только обозначениями от уравнения Аррениуса:
S = CAZ, где S - число видов, A - площадь, а C и Z - безразмерные
параметры. Предполагается, что наклон графиков, который определяется вели-чиной
Z, для островов будет более крутым, чем для материковых участков, а диапазон
значений Z для изолятов (островов) будет примерно равен 0.18-0.35, а для
выборок - 0.12-0.17 (Preston, 1962; MacArtur, Wilson, 1967). Теория была
подтверждена на многих примерах (Уилкокс, 1983). Не вдаваясь в подробный
анализ этих соотношений, отметим, что в Северной Охотии значение Z для
ост-ровов (0.133) меньше, чем для побережья (0.173), что прямо противоречит
тео-рии островной экологии. Этот факт можно интерпретировать как дополнитель-ное
доказательство несамостоятельности островных флор Северной Охотии. Другими
словами, флоры островов проявляют себя как выборки по отношению к флоре
побережья и, в строгом смысле, не являются изолятами. Без сомнения, эффект
изоляции начинает проявляться при удаленности от побережья боль-шей, чем
это имеет место быть в Северной Охотии (максимально удален от по-бережья
о-в Завьялова - на 19 км).
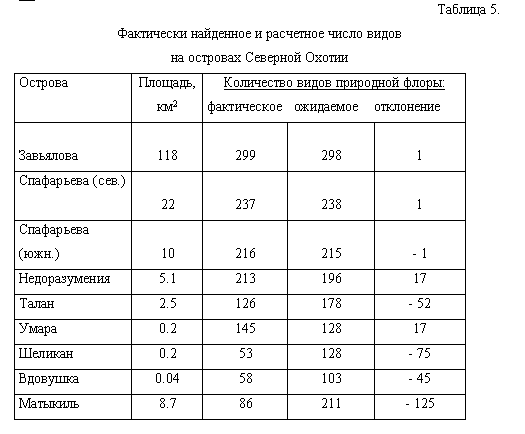
Если экстраполировать зависимость между числом видов и площадью острова,
установленную для о-вов Завьялова и Спафарьева (ОФ 1-3), на другие острова
Северной Охотии, можно вычислить разницу между фактическим и ожидаемым
количеством видов (табл. 5). Если ожидаемое количество видов больше, чем
фактическое, отклонение является величиной отрицательной. От-метим, что
мы не использовали для расчета тренд Вдовушка-Завьялова (линию регрессии,
соединяющую на рис. 6 островные флоры 8 и 1), поскольку в этом случае уравнение
линейной регрессии опирается даже не на три, как в преды-дущем случае,
а только на две, притом крайне неравнозначные островные фло-ры
Наибольшее отклонение от расчетного количества видов в сторону уменьшения
наблюдается на островах, флора которых обеднена в результате орнитогенного
воздействия (Матыкиль, Шеликан, Талан), а также на о-ве Вдо-вушка, видимо,
потому, что полученные нами постоянные a и z в уравнении Аррениуса нельзя
экстраполировать на столь ничтожную площадь. Не исключе-но также, что это
отчасти результат негативного влияния антропогенного фак-тора на уровень
видового богатства флоры о-ва Вдовушка (рекреационная на-грузка, пожары).
Большая величина отклонения для о-ва Матыкиль связана с неполной инвентаризацией
флоры: недоучет, однако, вряд ли составляет боль-ше 50 видов. Отклонения
в большую сторону отмечены для о-вов Умара и Не-доразумения, что можно
объяснить большей близостью их к побережью, но ка-кие конкретно факторы
имеют значение - лучшая защищенность от морских ветров, исторические причины
(обусловившие высокий уровень богатства фло-ры на противолежащих участках
побережья) или меньшая величина “островно-го эффекта” - остается неясным.
Этими общими рассуждениями о влиянии изо-ляции на уровень видового богатства
мы и ограничимся, потому что какой-либо математической зависимости между
числом видов на островах Северной Охо-тии и удаленностью их от побережья
не наблюдается.
На основании установленной нами корреляции между размером островов
и количеством обитающих на них видов можно делать выводы об относительной
бедности или относительном богатстве исследованных островных флор (в сравнении
с расчетной величиной), которые вполне согласуются с интуитивной интерпретацией
полевых наблюдений. Наиболее значимыми факторами, опре-деляющими видовое
богатство на островах Северной Охотии, являются размер острова и орнитогенное
воздействие, отмечено также относительное обогаще-ние флоры островов, близких
к участкам побережья с богатой флорой (о-в Умара - п-ов Кони, о-в Недоразумения
- устье р. Окса).
Кроме объективных факторов, влияющих на количественный и качест-венный
состав островных флор, нельзя не упомянуть о важности сведения к минимуму
факторов субъективных, особенно в связи с тем, что флора островов Северной
Охотии изучалась первоначально разными исследователями. На-сколько возможно,
мы постарались освободить анализируемый список от видов, определенных ошибочно,
от дублирующих синонимов и от различий, связанных с толкованием границ
таксонов. При любых количественных сравнениях важно также, чтобы степень
флористической изученности сравниваемых единиц была достаточно высокой
и равномерной. Известны высказывания (Connor, Simberloff, 1978), что число
видов в списке в большей степени зависит от числа ботанических экспедиций,
побывавших на острове, нежели от его площади, вы-соты над уровнем моря
или изоляции. Отметим, что в результате тщательных и целенаправленных полевых
исследований флора островов Тауйской губы выяв-лена на 95-99%. Наилучшим
образом обследованы небольшие острова, а наи-большего пополнения списка
следует ожидать для о-ва Завьялова, где некото-рая часть территории осталась
неизученной из-за труднодоступности. С учетом большой нивелирующей роли
орнитогенного влияния на условия обитания рас-тений и связанного с этим
снижения флористического разнообразия, изученность флоры о-ва Матыкиль
(86 видов) составляет не менее 70%.
назад